This work has been funded by the Yale Institute for Biospheric Studies and the National Science Foundation.
The pioneering work of Kölreuter and Sprengel in the mid- to late-1700s established that flowers were–contrary to popular opinion of the time–involved in sexual reproduction. This discovery established the nascent field of pollination biology that has shaped most of what we know today about floral evolution.

Sprengel’s pioneering 1793 work on plants and pollinators.
Yet, mounting evidence over recent decades has shown that other agents of selection inbesides only pollinators can be important drivers of floral trait evolution. Indeed, flowers must perform multiple functions, from attracting pollinators to protecting the developing embryo. The importance of these functions can differ between the sexes: only female flowers must protect the developing emrbyo. Furthermore, the non-pollinator agents of selection, which include the physiological costs of flower production and maintenance, can exert stronger selection on flowers than biotic agents, both pollinators and enemies Caruso et al. 2018. Additionally, traits under selection by pollinators can affect flower physiology in complex ways (Roddy 2019, IJPS).
Physiological costs of flowers
Because non-pollinator agents of selection often oppose pollinator selection (e.g. pollinators prefer larger flowers, but larger flowers are more costly to produce), the physiological costs of maintaining and producing flowers represent a critical dimension of floral evolution. Yet, there has been surprisingly little work aimed at characterizing these costs in a comparative, phylogenetic framework.
The first comparative evidence suggests that early-divergent ANA-grade and magnoliid flowers are much more costly to produce than monocot and eudicot flowers, per unit display area. This suggests that early in angiosperm evolution the physiological costs of floral display was drastically reduced.
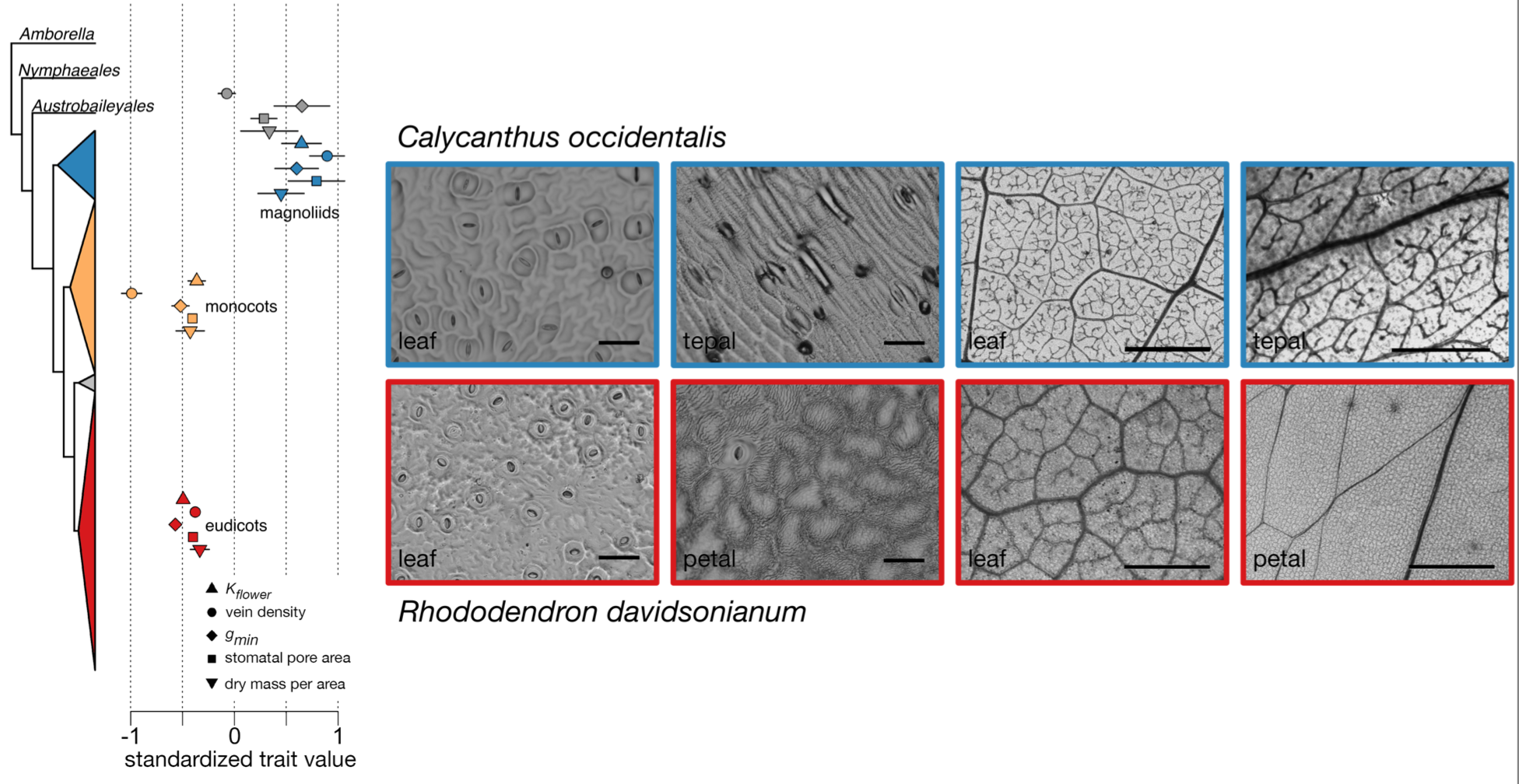
Compared to ANA grade and magnoliid flowers, monocot and eudicot flowers have fewer stomata, fewer veins, and are less costly per unit area in terms of both water and carbon (Roddy et al. 2016; Roddy 2019).
But how cheap can flowers be? For floral display structures, reducing the physiological costs per unit display area may be beneficial, but the organ must remain upright and on display. Because carbon is metabolically expensive, flowers may rely on a turgor-driven hydrostatic skeleton to remain upright. Consistent with this hypothesis, flowers have significantly higher water contents than leaves (Roddy et al. 2019), which allows them to maintain high water potentials despite large declines in water content.
Further evidence that flowers rely on a hydrostatic skeleton to remain upright comes from microCT imaging of desiccating Calycanthus flowers, which revealed that after turgor loss most of the air space in floral tepals (analogous to petals) disappeared (Roddy et al. 2018). In other words, without intracellular turgor the 3D tissue structure collapsed, likely because there is so little biomass invested to maintain the structure upright.
This idea is being explored more fully using microCT imaging and in collaboration with soft matter physicists to characterize and model tissue structure and mechanics.
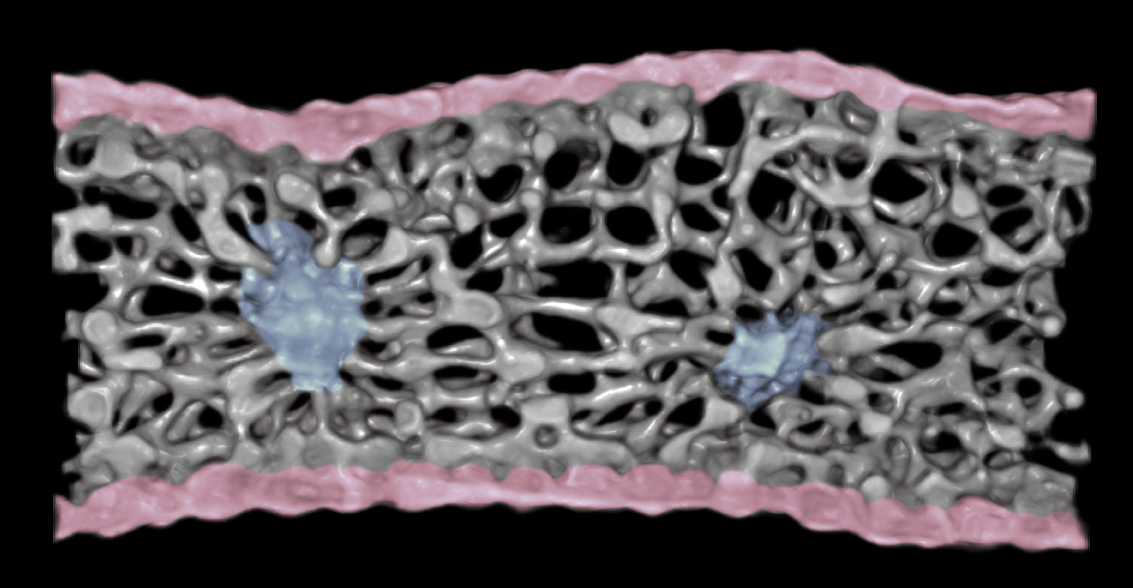
3D microCT image of fresh and hydrated Rhododendron cilipes (Ericaceae) petal. Epidermal cells are colored pink and veins are colored blue. The mesophyll cells form a meshlike network.
Modularity and ecology
Like leaves, flowers develop from the shoot meristem and are borne on aerial shoots. Yet flowers are heterotrophic, relatively short-lived, and must perform different functions. The extent to which traits in flowers and leaves are correlated (i.e. how modular are flowers and leaves; Roddy et al. 2013; Roddy et al. 2019) will determine (1) how selection on one organ may translate into selection on the other and (2) how habitat filtering for, say, vegetative traits may influence the distribution of floral traits in a community. One current project, led by PhD student Dario Caminha Paiva, is sampling floral traits along an elevational gradient in the campo rupestre ecosystem of Brazil.
One hindrance to quantifing the modularity of the plant bauplan has been that the same traits are rarely measured on both flowers and leaves. Yet, measuring the same traits in multiple organs clarifies which traits and trait relationships are fundamental to building a structure and in which dimensions phenotypes are free to roam. Effectively, such studies can highlight the role of selection and the importance of adaptation (Olson and Pittermann 2019) in driving trait relationships.
A whole-plant framework that unifies our often disparate knowledge of reproductive and vegetative structures will enrich our understanding of the multiple axes of selection that have shaped plant form and function and that drive ecological distributions.
Flowers in a changing climate
Successful reproduction depends on the flower’s ability to remain physiologically functional. Yet, climate anomalies introduce novel selective regimes that may alter the ability of a flower to remain viable and functional. At the same time, animal pollinators are declining in abundance, which would increase competition among flowers for limited pollinators, potentially driving plants to invest more resources in reproduction. Extreme temperatures and droughts reduce resource avaiability and impose physiological challenges to supporting flowers. Together, these lines of evidence suggest that the physiological costs and tolerances of flowers may turn out to be critical in determining which species may survive the rapidly shifting climates that are altering selection on flowers.
Related papers
Roddy et al. 2021. Towards the flower economics spectrum New Phytologist
Roddy 2019. Energy balance implications of floral traits involved in pollinator attraction and water balance. International Journal of Plant Sciences
Roddy et al. 2019. Hydraulic traits are more diverse in flowers than in leaves. New Phytologist
Roddy et al. 2018. Water relations of Calycanthus flowers: Hydraulic conductance, capacitance, and embolism resistance. Plant, Cell & Environment
Roddy et al. 2016. Hydraulic conductance and the maintenance of water balance in flowers. Plant, Cell & Environment
Roddy et al. 2013. Uncorrelated evolution of leaf and petal venation patterns across the angiosperm phylogeny. Journal of Experimental Botany
Roddy and Dawson 2012. Determining the water dynamics of flowering using miniature sap flow sensors. Acta Horticulturae